

Freshwater ecosystems are crucial hotspots of biodiversity, yet their underwater landscapes often remain hidden from our view, presenting significant challenges for researchers and conservationists. In Croatia’s stunning Dalmatia, the intricate and deep karstic lakes posed a particularly complex puzzle for habitat mapping and biodiversity assessment. Traditional methods, reliant on point sampling from boats or visual census through diving, provided only fragmented insights at high costs, rendering comprehensive mapping and monitoring near impossible.
Enter a multidisciplinary endeavor led by BIOTA Ltd, employing cutting-edge technology and innovative approaches to unveil the mysteries of the Dalmatian lakes’ transitional habitats and biodiversity. At the heart of this project lay the utilization of side-scan multi-beam sonars, a technology often underutilized in civilian applications but offering immense potential for scientific exploration.

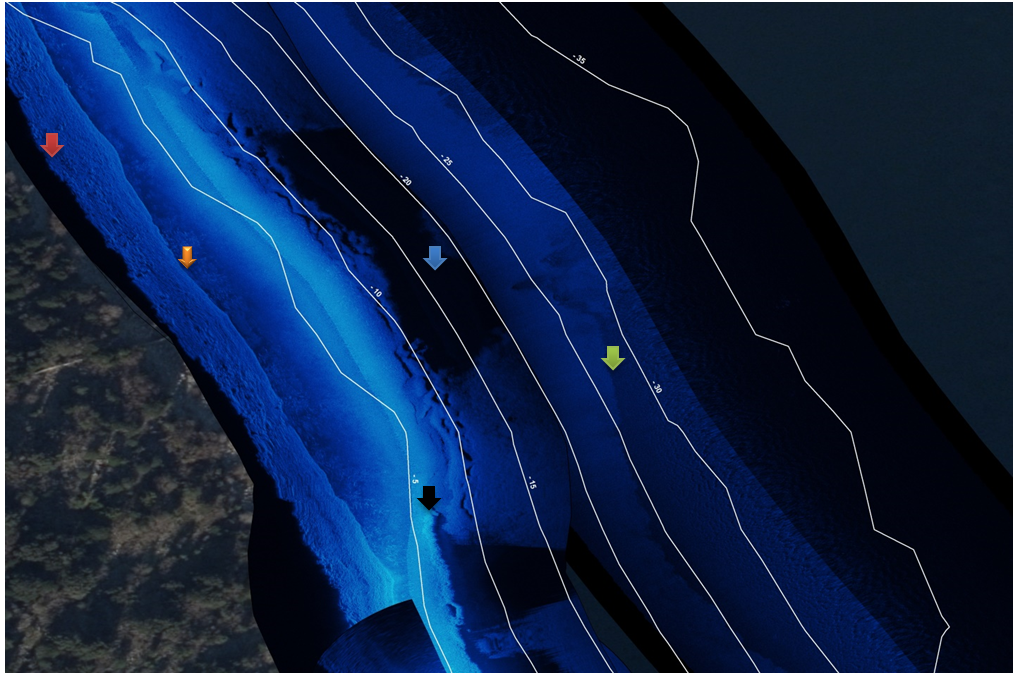
Our expert team embarked on an intensive field campaign, meticulously scanning the depths of different Dalmatian lakes. Mounted on a specialized aluminum boat equipped with state-of-the-art sonar equipment, we navigated predetermined recording lines, capturing detailed imagery of the lake bed and its inhabitants. The result? A wealth of data revealing not just the contours of the lake bottom, but also the distribution of vegetation, substrate types, and even the presence of wildlife, all with unprecedented clarity and precision.
But data collection was just the beginning. Leveraging Geographic Information System (GIS) technology, we translated raw sonar images into comprehensive maps, delineating habitat types and their extent with remarkable accuracy. From dense vegetation patches to rocky outcrops, every feature beneath the water’s surface was meticulously documented, providing invaluable insights into the complex ecosystems thriving within these lakes.
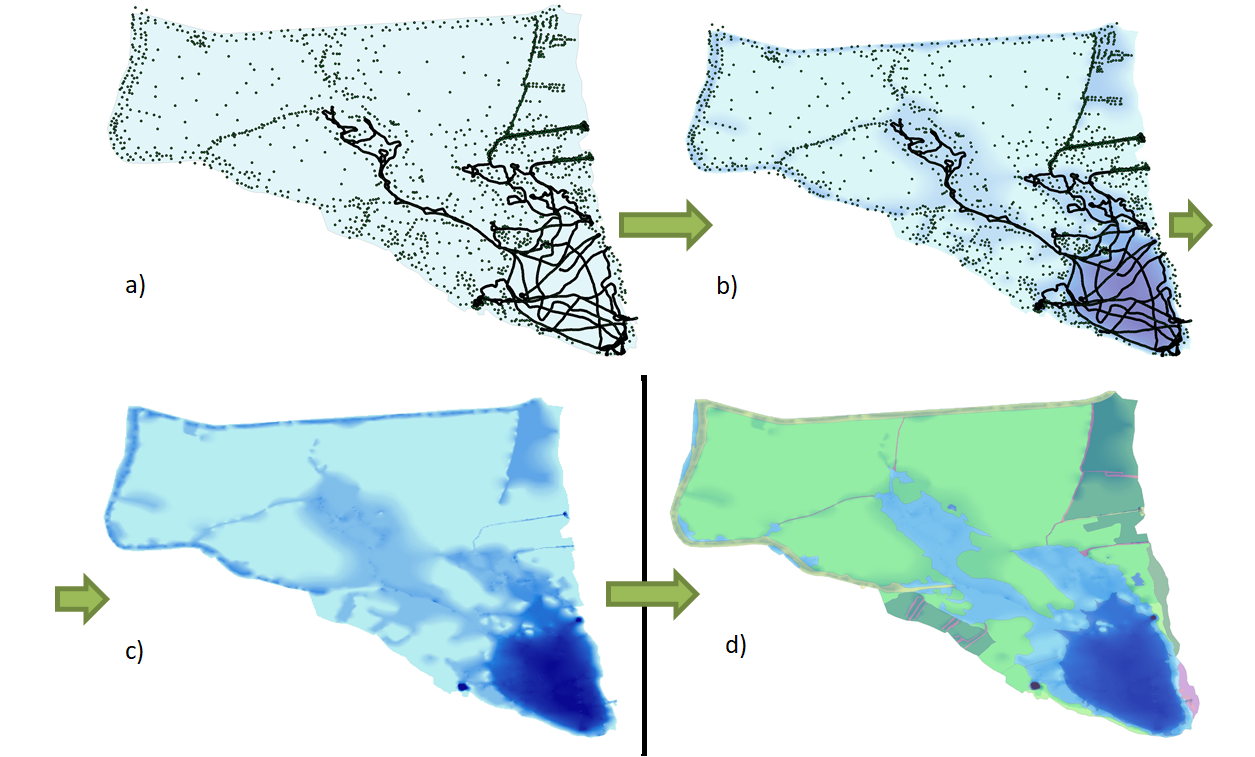
However, our approach didn’t stop at remote sensing. Recognizing the importance of ground truthing, we complemented our sonar surveys with meticulous vegetation sampling, employing a combination of diving expeditions and innovative underwater filming techniques. By collecting vegetation samples and visually documenting submerged habitats, we not only validated our remote sensing data but also gained deeper insights into the composition and dynamics of these fragile ecosystems.
Central to our mapping efforts was the integration of established habitat classification systems, such as the Croatian National Habitat Classification (NKS). By aligning our findings with existing frameworks, we ensured compatibility and comparability with broader conservation initiatives, facilitating informed decision-making and policy development.
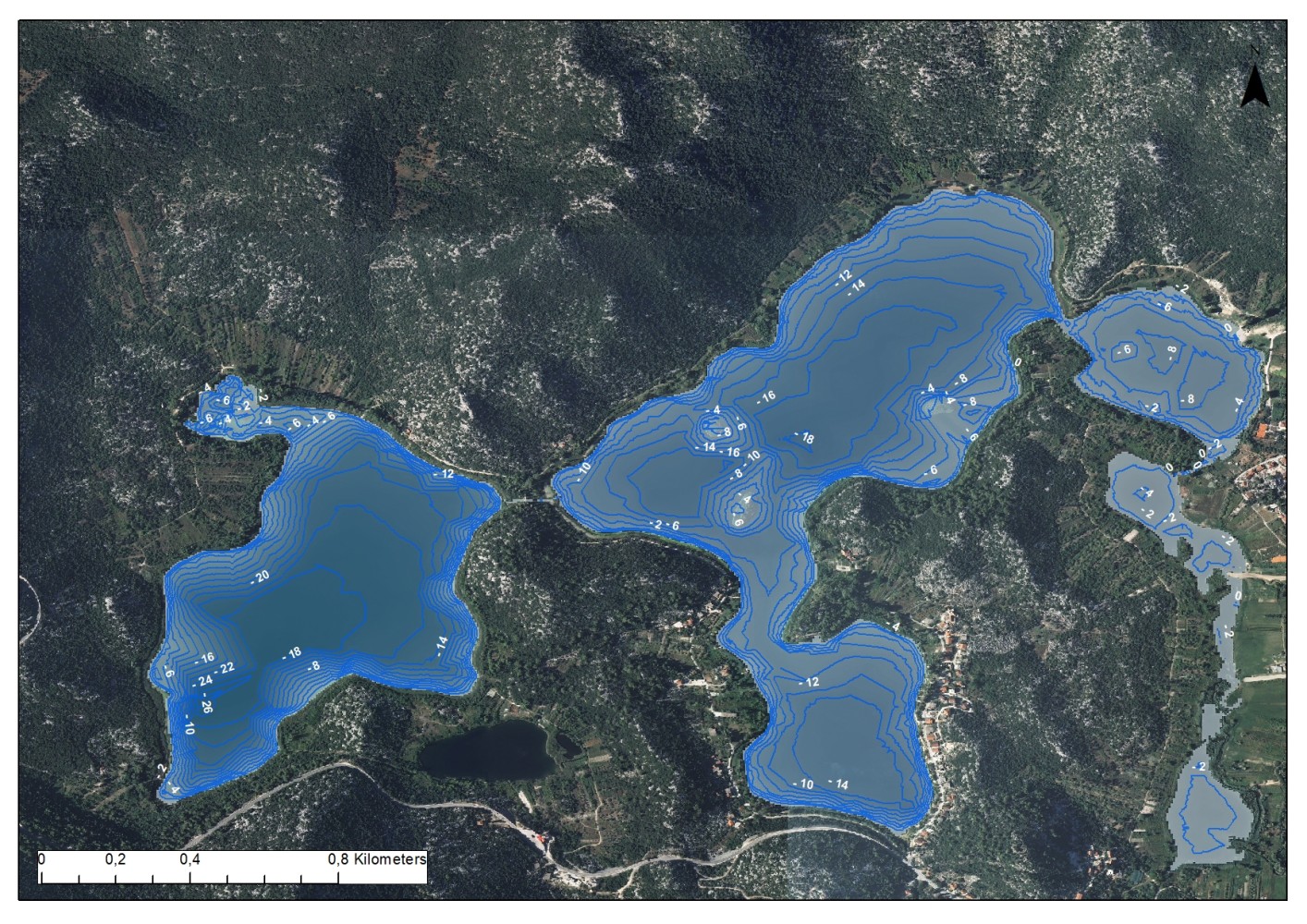
Habitat data was filled in with data from transects and point counts done by diving. Diving was done during high freshwater vegetation season and transects were set to follow the coast slope (from water edge to deepest point). Vegetation is mainly only found in light penetration zone up to – 15 m, but dives were done up to the max -45 m.

Yet, our work goes beyond mere cartography. It’s about understanding and safeguarding the delicate balance of life within the lakes. By identifying and assessing potential threats to habitat integrity, from invasive species to anthropogenic impacts, we’re paving the way for targeted conservation strategies aimed at preserving these invaluable ecosystems for generations to come.

In essence, our project represents a paradigm shift in freshwater habitat mapping, demonstrating how innovative technologies and collaborative approaches can unlock the secrets of hidden ecosystems. As we continue to refine our methodologies and expand our reach, we’re not just mapping habitats; we’re illuminating the pathways to a more sustainable future for our planet’s precious freshwater resources. Join us on this journey of discovery, where every dive into the depths brings us closer to understanding and protecting the wonders that lie beneath.
This type of habitat mapping can also be applied to rivers and streams. If you are interested in this service in South and Central Europe, feel free to contact us.




0 Comments